
12 Startups Applying AI to Gene Editing: From Custom CRISPR to Zinc-Finger Revivals
Despite only 1% of biotech VC going to gene editing in 2024, a wave of AI-driven startups is advancing along CRISPR’s clinical milestones


In May 2025, clinicians at Children’s Hospital of Philadelphia and Penn Medicine successfully performed a personalized CRISPR-based therapy on a newborn diagnosed with CPS1 deficiency—a rare genetic disorder impairing ammonia clearance in the liver. Built on an adenine base-editing platform, the treatment was developed and administered within six months of diagnosis, stabilizing the infant’s condition and marking reportedly the first known case of a bespoke in vivo gene-editing intervention in a single patient.
While the CHOP–Penn therapy was built on established molecular and delivery technologies, it highlighted the pace that targeted interventions can now reach. For many, the next question is how such approaches might scale beyond bespoke development. One path involves increasing reliance on computational tools to guide design and reduce iteration time. Reflecting on that broader possibility in late 2024, CRISPR co-inventor Jennifer Doudna noted:
“It’s still early days, but the potential to appropriately harness the joint power of AI and CRISPR, arguably the two most profound technologies of our time, is clear and exciting—and it’s already started.”
In this article: The Cradle of Gene Editing—CRISPR Arrives—Life after CRISPR—AI-Powered Applications—Startups Applying AI—DNA Cleavage Layer—DNA-Base Editing — RNA Editing—Epigenetic Rewiring—CRISPR-Customising Layer—Policy, Funding, and the Future
The Cradle of Gene Editing
Genetic engineering began in 1974 with an experiment in mice, where Rudolf Jaenisch and Beatrice Mintz injected viral DNA into early embryos and showed it had integrated into the animals’ germline. In 1980, Jon Gordon and Frank Ruddle advanced the method by microinjecting foreign DNA into fertilized eggs, producing mice that passed the genetic changes to their offspring. The term “transgenic” was introduced a year later to describe these genetically altered animals.
“Important as transgenic mice are, they are really the tip of the iceberg when compared with what we are going to see in the next few years.”—Janet Rossant (National Research Council, 1994)
In 1981, two groups—Martin Evans with Matthew Kaufman, and independently Gail Martin—isolated pluripotent embryonic stem (ES) cells from mouse blastocysts. This made way for targeted gene deactivation using homologous recombination (a method that replaces a stretch of DNA with an engineered copy by using the cell’s own repair machinery) in ES cells. In the late 1980s, Oliver Smithies and Mario Capecchi independently applied that technique, and together with Evans, shared the 2007 Nobel Prize in Physiology or Medicine.
Shortly after, the Cre-lox system, originally derived from a bacteriophage, introduced temporal and spatial control to genetic alteration (pioneered by Nat Sternberg, and later adapted for mammals by Brian Sauer and Nancy Henderson).
A marked advance came when researchers showed, first with the yeast-derived meganuclease, that introducing a site-specific double-strand break (DSB) in mammalian DNA massively increases the cell’s uptake of foreign sequences.
These nucleases (enzymes that cut DNA) trigger the cell’s repair machinery, which can act in two ways:
Error-prone non-homologous end joining (NHEJ), leaving random insertions and deletions (INDELs).
High-fidelity homology-directed repair (HDR) when a suitable donor template is present, enabling precise sequence replacement or gene insertion.
Although meganucleases such as I-SceI showed the concept works, each one cuts only a single 20–30-bp DNA sequence, so scientists had to scrupulously redesign the enzyme for every new target, making it clear we needed nucleases that can be retargeted far more easily.
In 1996, scientists built zinc-finger nucleases (ZFNs) by attaching a customizable DNA-binding module (“handle”) to Fok1, a nuclease derived from a marine bacterium. By 2005, this breakthrough had sparked the phrase “genome editing”, as ZFNs let researchers tweak genes with pinpoint accuracy, even turning off specific genes in animals like rats.
In 2010, the discovery of transcription activator–like effector nucleases (TALENs) offered more flexible DNA-binding modules in comparison to ZFN further simplifying genome editing. Together, these three—meganucleases, ZFNs, and TALENs—laid the groundwork for today’s advanced gene-editing.
CRISPR Arrives
Imagine a nuclease merging the precision-binding prowess of ZFNs and TALENs, but sidestepping all the painstaking protein engineering, because nature had already perfected the mechanism.
In the late 1980s, curious genetic patterns were discovered in E. coli genomes: clusters of short DNA repeats, each separated by unique sequences that didn’t repeat. The purpose of these alternating repeat–spacer patterns was unknown at the time.By the early 2000s, scientists realized these arrays, now called Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR), were consistently present with Cas genes across bacteria and archaea.
By 2007, Horvath’s group uncovered CRISPR/Cas as microbial adaptive immunity, a molecular memory bank storing viral genetic snippets (spacers) to guide precision-strike nucleases that recognize and cut matching viral sequences during future infections. Central to this system is the CRISPR RNA (crRNA), directing the nuclease, and the protospacer-adjacent motif (PAM), specifying the exact genomic target.
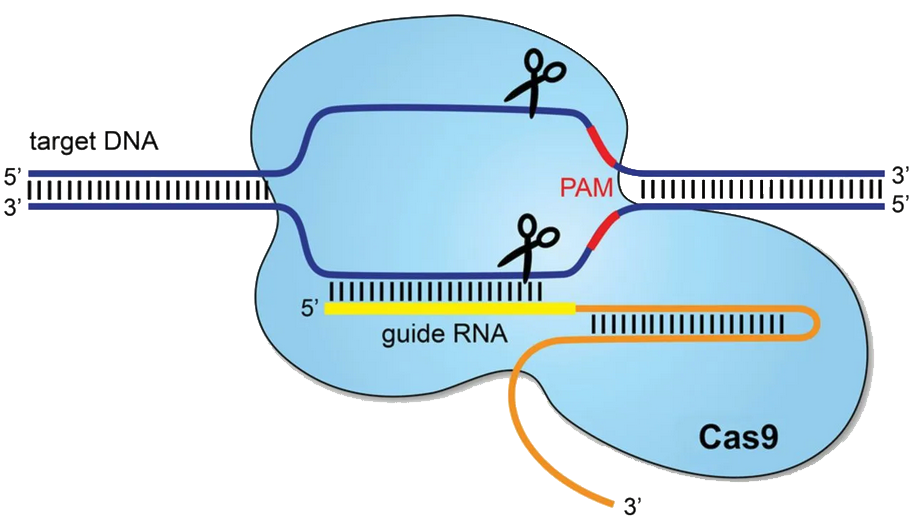
In 2012, at Berkeley and Umeå, Emmanuelle Charpentier and Jennifer Doudna adapted this natural microbial defense, reprogramming Cas9 nuclease by fusing crRNA and trans-activating crRNA into a streamlined single-guide RNA. Within a single lab session, they converted bacterial immunity into a universal genome-editing scalpel. Just months later, Feng Zhang and George Church introduced this innovation into mammalian cells, easily achieving genome modifications without the need for intricate protein redesign. This earned Doudna and Charpentier the Nobel Prize in Chemistry in 2020.
For a fresh visual overview of CRISPR-based tools (including the emerging bridge RNA), see: A Visual Guide to Genome Editors by Evan DeTurk and Ella Watkins-Dulaney (Asimov Press, 2025).
Life after CRISPR
While it began as a bacterial “molecular scissors” called Cas9 that could cut DNA at almost any chosen spot, the original CRISPR was bulky and sometimes sliced the wrong place. Scientists quickly built smaller versions that fit inside common gene-therapy delivery viruses, and they discovered sister enzymes like Cas12a, which can reach DNA regions the first Cas9 missed, and Cas13, which edits RNA instead of DNA for temporary effects.
Meanwhile, protein-engineering tweaks have made the scissors far more accurate, reducing the risk of unwanted genetic changes. Newer CRISPR spin-offs go a step further:
base editors swap single DNA letters without cutting the strand at all;
prime editors, discovered in 2019, can write short custom edits (additions, deletions or replacements) with considerable precision.
Even when the cutting action is disabled, CRISPR still serves as a programmable “GPS” that can ferry in other molecules to switch genes on or off or illuminate chromosomes for imaging.
AI-Powered Applications
As the field advances, it’s increasingly paired with AI and pharmacogenomics to guide therapy design. A 2025 Pharmaceutics review highlights DeepSpCas9, a deep-learning model (trained on >12,000 guide sites) that predicts cut efficiency before an experiment. Its alternative CRISPRon is another DL tool that predicts how often Cas9 will introduce edits at the intended DNA site.
When combined with patient-specific pharmacogenomic data, these AI tools help guide drug selection and dosing. In parallel, multiplex CRISPR editing enables off-the-shelf CAR-T therapies like CTX110, where T cells are engineered to remove their native receptors—avoiding graft-versus-host complications—and redirected to target relapsed B-cell cancers.
In 2023, ex-vivo gene therapy Casgevy (Vertex/CRISPR Therapeutics) became the first CRISPR-based medicine to receive FDA approval, marking a clinical milestone in treating sickle-cell disease (back in 2019, Victoria Gray was the first patient).
Since then, development has advanced across three main areas:
Medicine: Intellia’s NTLA-2002 have reached Phase III, Verve’s cholesterol-lowering VERVE-102 programme successfully completed Phase 1b, Prime Medicine’s PM359 introduced prime editing to patients, and Excision’s EBT-101 for HIV completed initial dosing. In early 2025 Tune Therapeutics raised $175M to develop Tune-401 for Hepatitis B.
Agriculture: gene-edited mustard greens by Pairwise are already on U.S. shelves, with seedless blackberries, stay-green bananas and vitamin-D-enhanced tomatoes being in development.
Biodiversity: Colossal Biosciences announced on 8 April 2025 the birth of three dire wolf-like pups engineered with 20 genomic edits drawn from 13,000- and 72,000-year-old dire-wolf DNA.
Behind these advances is a growing reliance on AI to optimize nearly every layer of genome editing, from target selection and guide design to protein engineering and delivery. Next we examine how computational tools are applied to each step of this process.